
2023年8月30日,艾沐蒽助力陆军军医大学在《细胞与分子免疫学》(Cellular & Molecular Immunology)(IF=24)上发表题为“衰老骨髓造血干细胞劫持调节性T细胞营造促其存活的微环境(Aged hematopoietic stem cells entrap regulatory T cells to create a prosurvival microenvironment)”的研究论文。研究使用艾沐蒽生物ImmuHub® TCR分析系统,研究解析了电离辐射致骨髓造血远期损害的效应与机制,拓展了衰老造血干细胞(HSCs)存活及优势扩增发生机制的认识,为电离辐射致骨髓造血远期损害的防治提供了新思路。
下面就让我们一起了解一下!

研究背景
组织驻留体细胞稳态的解除被认为是衰老过程的关键因素,造血干细胞(HSCs)的衰老支撑着系统器官有限的健康跨度,造血干细胞是研究干细胞衰老的一个很好的范例。HSC老化的特征是重构能力降低、骨髓偏向性HSCs扩大、复制应激增加、DNA突变积累、细胞凋亡抵抗能力增强、自噬能力丧失以及表观遗传、转录和翻译改变。然而,HSC老化这些特征的机制相互关系和等级顺序尚未明确。特别是,引起HSC衰老的起始事件的性质和时间在很大程度上仍然未知,因此无法从早期预防HSC衰老。
该研究使用早衰和中年小鼠模型,发现并表征了BM Tregs在HSC衰老中的独特功能。研究发现,衰老HSCs中的DNA突变积累可诱导主要组织相容性复合体II类(MHCII)上调和BM Treg克隆扩增。此外,通过MHCII -T细胞受体(TCR)的参与,衰老HSCs与BM Tregs之间建立了双向相互作用,通过TCR识别衰老HSCs上的MHCII,BM Tregs得以扩展,而通过间隙连接(GJ)介导的BM Tregs中环磷酸腺苷(cAMP)的转移,衰老HSCs的凋亡启动得以减弱。重要的是,靶向HSC-Tregs相互作用可以防止HSC在早期衰老。
方法学
-
RNA-seq:在这项研究中,使用流式细胞术对于HSC和BM细胞进行细胞分选,从小鼠骨髓中分选造血干细胞和treg,提取RNA,进行RNA-seq。
-
TCRβ扩增和测序:TCRβ测序时,对BM Tregs进行分类,并提取RNA。随后,通过艾沐蒽生物 ImmuHub®技术对BM Tregs TCRβ链采用5 ‘ RACE法进行文库构建和高通量测序。
-
全外显子组检测:将5只小鼠的骨髓造血干细胞进行分类汇总,提取基因组DNA进行深度WES检测。
研究结果
01 HSC早期衰老伴随BM Treg克隆性扩增
最初,作者采用了一种公认的小鼠早衰模型,将小鼠暴露于电离辐射(IR)中,这是一种众所周知的诱变剂。这与生理性造血老化类似,HSC老化的特征,包括扩张(图1A),重构能力受损(图1B)和骨髓偏向分化(图1C)是明显的,在IR (mpi)后3个月小鼠中明显可见,同时BM细胞数量适度增加(补充图1b)。在转录组水平上,与老年小鼠(18-24个月大)类似,IR小鼠的BM HSC表现出髓系偏倚(图1D),并且使用aging signature(图1E)交叉验证了衰老特征,该特征提供了小鼠HSC衰老的全面转录组特征。令人惊讶的是,对于BM T细胞,作者观察到CD4+ T细胞在IR小鼠中扩增(图1F,G)。在CD4+T细胞中,除常规CD4+T细胞(Tconv)外,Foxp3+ Tregs的比例和数量在IR小鼠中显著增加(图F,G)。大多数位于外周组织的调节性T细胞是从胸腺输出的(称为tTreg)。这与生理性衰老类似,IR小鼠胸腺细胞,包括Treg数量,由于IR诱导的胸腺萎缩显著下降(图1H)。Treg的积累也可能源于Tconv的Treg表型转化(称为pTreg)或组织内Treg的局部扩增。根据使用这些pTreg生成的转录组数据,作者将pTreg特征叠加到火山图上,并将IR小鼠的BM Treg转录组与对照(Ctrl)小鼠的转录组进行比较,发现没有明显的Tconv-Treg表型转换迹象(图1I)。与脾Tregs (SP)和淋巴结Tregs (LN)相似,BM Tregs具有HelioshiNrp-1hi表型(图1J),这被认为是区分tTregs和pTregs的一种表型。这些数据表明pTregs对IR后BM Treg扩张的贡献很小。TCR库显示IR小鼠中BM Tregs的克隆扩增(图1K, L)。此外,作者还发现IR小鼠的BM中Ki67+ Tregs的比例显著增加,而不是SP或LN(图1M),反映了BM Tregs的特异性克隆增殖。
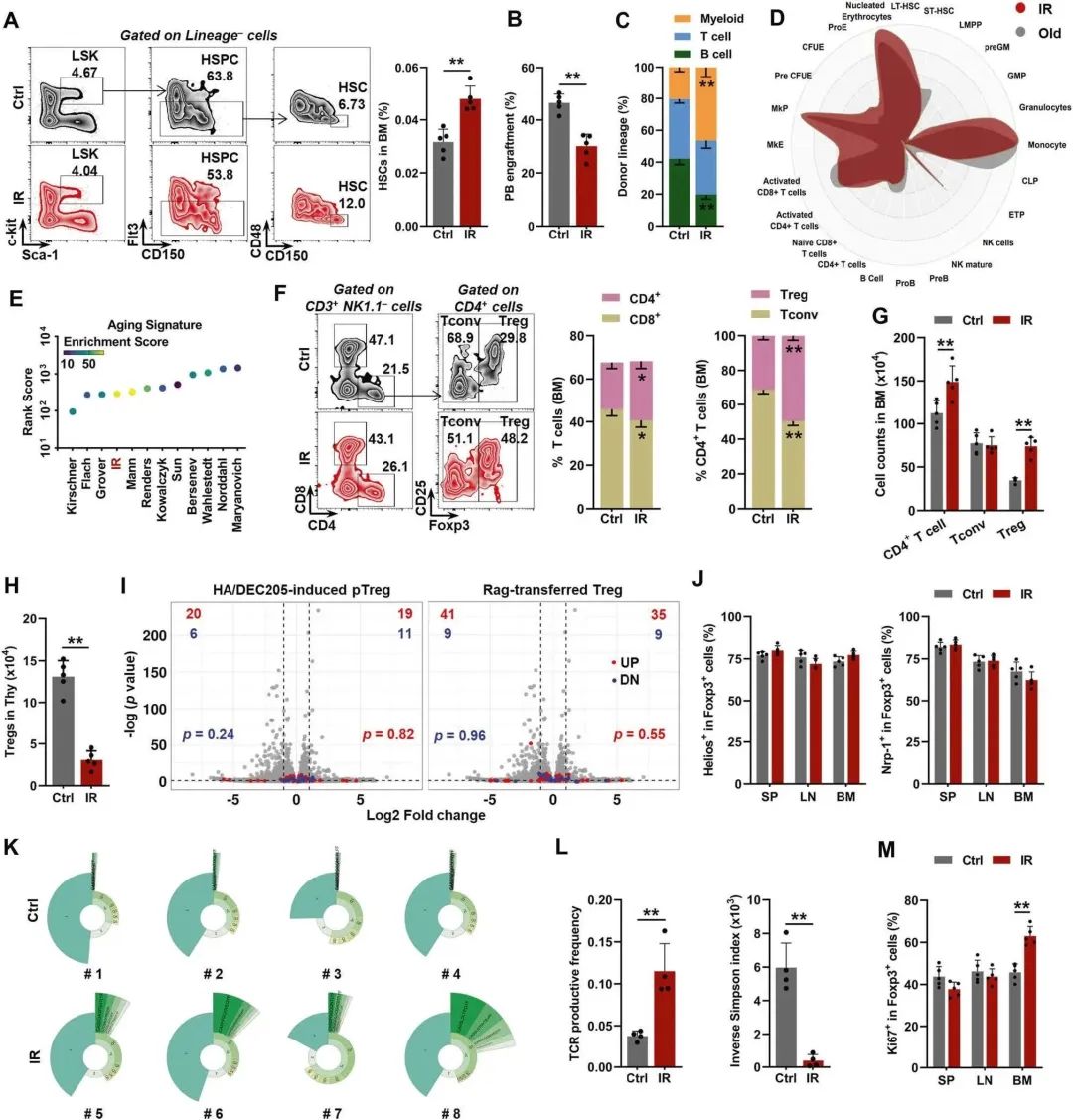
图1
02 MHCII-TCR参与触发老年HSC和BM treg之间的双向相互作用
BM Tregs的克隆扩增表明,BM Tregs可能通过TCR识别BM驻留抗原呈递细胞(APCs)表面的MHCII呈递的同源抗原而被激活。实际上,通过RNA-seq观察到IR小鼠BM Tregs中TCR信号的显著激活(图2A)。通过分析了MHCII在骨髓中的专业APCs(包括Dc细胞、B细胞和巨噬细胞(MACs))以及最近被鉴定为熟练APCs的HSC上的表达。在IR小鼠的HSC中检测到MHCII而非MHCI的显著上调表达(图2B)。相应地,RNA-seq也揭示IR小鼠造血干细胞中MHCII介导的抗原呈递途径明显激活(图2A)。鉴于BM Tregs在人体和小鼠体内稳态时都定位于HSC附近,MHCII在IR小鼠的HSC上呈献同源抗原可能是BM Tregs克隆扩增的原因。为了支持这一发现,组织学分析显示,与对照组相比IR小鼠的造血干细胞,特别是MHCII+造血干细胞,比骨髓中的Tconv或CD8+T细胞更接近Treg(图2C-E),47.2%的造血干细胞与直接的Treg接触(在Treg10μm内)。根据MHCII的表达,作者将造血干细胞划分为MHCIIhi和MHCIIlo亚群(图2F)。作者发现,从1 mpi开始,MHCIIhi HSC的比例和数量逐渐急剧增加,而MHCIIlo HSC的比例急剧减少,尽管它们的数量只是轻微下降(图2G)。在与OTII naïve CD4+ T细胞共培养中,证实来自IR小鼠的MHCIIhi HSC比它们的MHCIIlo同类具有更强的抗原呈递能力。在体外,在MHCIIhi HSC与IR小鼠自体的BM Treg共培养系统中,观察到显著的Treg增殖,而通过Transwell室分离或用单克隆抗体(αMHCII)阻断MHCII显著地消除了增殖作用(图2H)。这些数据表明,TCR对IR小鼠造血干细胞上MHCII的识别刺激了BM Tregs的克隆扩增。
主成分分析(PCA)显示,IR小鼠的MHCIIhi HSC与对照小鼠的MHCIIhi和MHCIIlo HSC存在很大差异,而IR小鼠的MHCIIlo HSC与对照小鼠的MHCIIhi和MHCIIlo HSC仅存在中度差异(图2I)。同时,IR小鼠的MHCIIhi HSC表现出更高的衰老特征等级评分(图2J)。这意味着MHCII上调可能标志着HSC过早衰老。为了支持这一发现,通过来自IR小鼠的MHCIIlo HSC相比,它们的MHCIIhi对应物在外周血(PB)和BM中都显示出明显较低的重建能力,并伴有更明显的骨髓偏倚(图2K)。然而,阻断MHCII或Transwell培养几乎可以消除这些影响(图2L)。在体内,Foxp3GFP-DTR小鼠的Treg消耗也显著挽救了MHCIIhi HSC的过度表达和HSC的老化表型(图2M, N),而不诱导Tconv和CD8+ T细胞的显著激活。过DT短期消耗Treg后Treg数量反弹在IR小鼠中仅部分发生,这可能是由于胸腺萎缩和激活的Tconvs缺乏Treg转化。特别是,相当比例的BM Tregs在稳态过程中定位于造血干细胞和CD8+ T细胞附近,CD8+ T细胞直接和/或间接调节正常和异常造血干细胞的维持。然而,CD8+ T细胞缺失对IR小鼠的HSC衰老表型或Treg缺失的治疗效果几乎没有影响。然而,BM Tregs的过继转移消除了这种拯救作用(图2M, N)。此外,转移的tregs占BM tregs的大部分,并且在受体IR小鼠中显示出显著的增殖(图2O)和在HSC生态位中的积累。总之,这些结果表明MHCII-TCR参与触发了衰老的HSC和BM treg之间的双向相互作用,这与驱动HSC衰老有关。
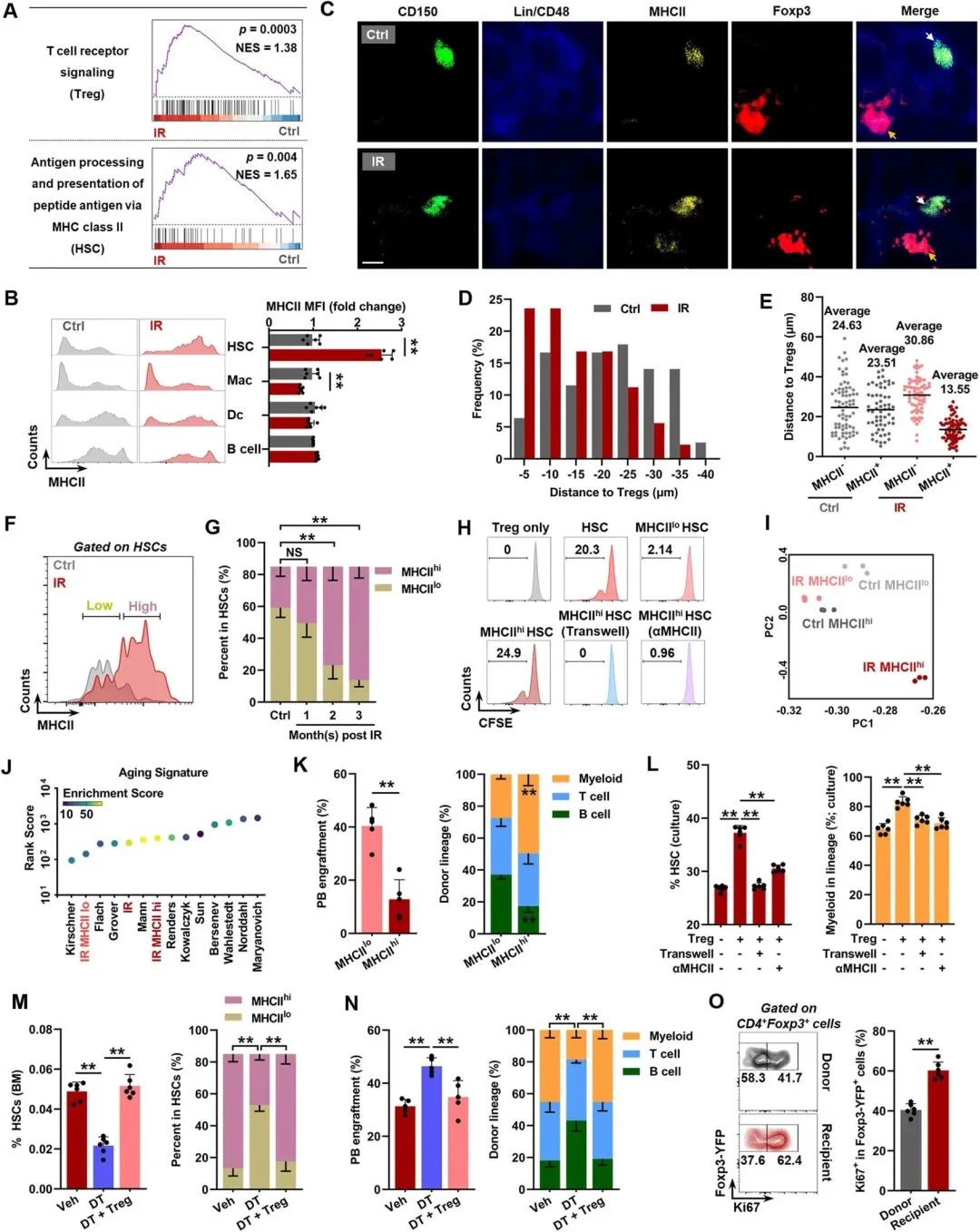
图2
03 DNA突变导致HSC MHC II上调
通过观察到IR小鼠的MHCIIhi HSC比MHCIIlo HSC表现出更显著的突变负荷,这是通过全外显子组测序(WES;图3A, B)。然而,当Tregs被耗尽时,IR小鼠造血干细胞中的DNA突变负荷显著减少,而BM Tregs的继发性转移几乎消除了这种影响(图3C, D)。这些数据表明,突变积累可能通过产生MHCII限制性新抗原来促进IR小鼠HSC中MHCII的上调,抗原特异性Tregs可能促进这些突变积累的MHCIIhi HSC的存活。
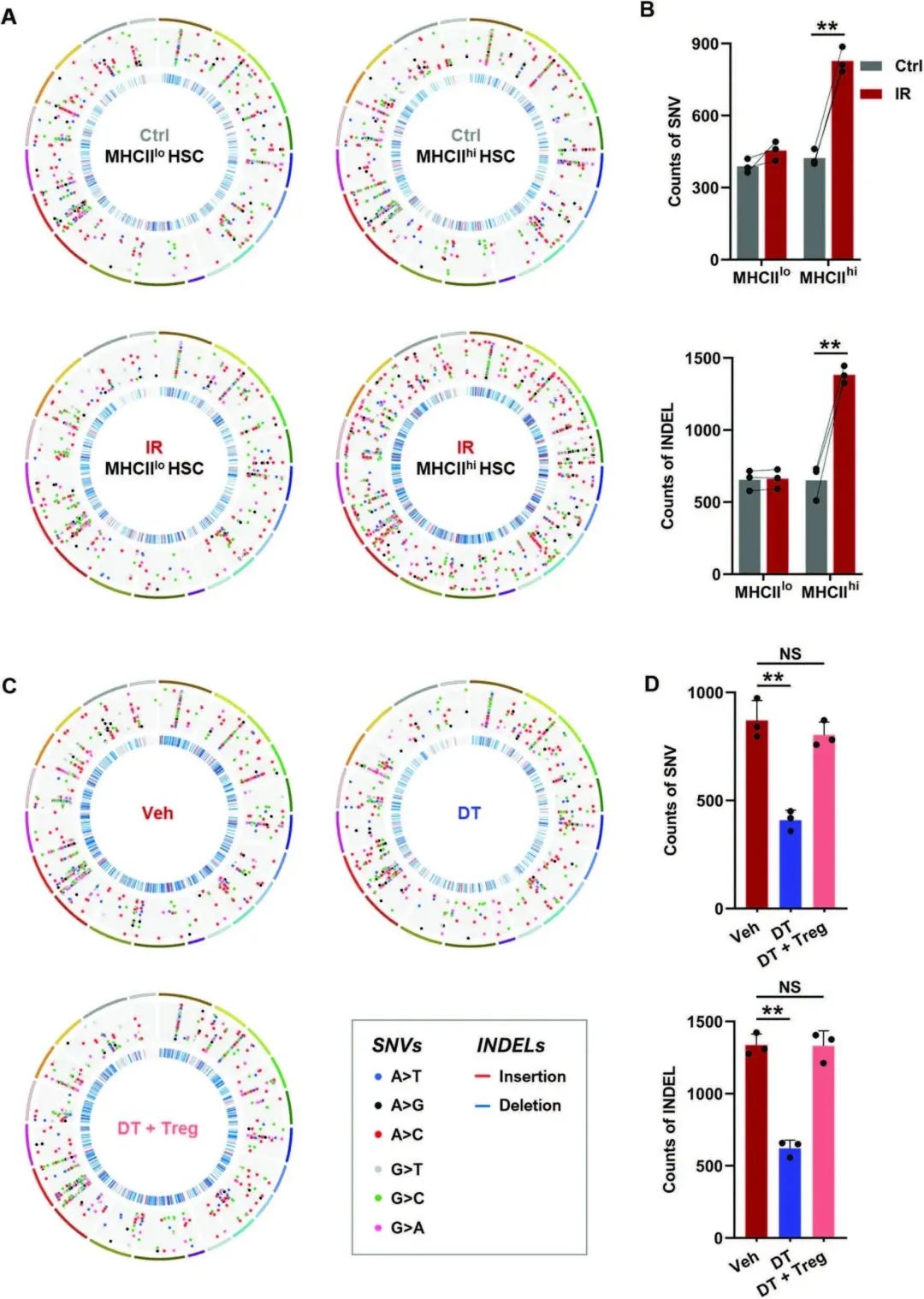
图3
04 衰老HSC通过Cx43与BM treg建立功能性GJs
接下来,作者试图确定MHCIIhi型HSC扩增的分子机制。首先分析了IPA对信号通路的最高富集,并注意到GJ信号在IR小鼠的BM HSC和Tregs中都被强烈上调(图4A)。据报道,MHCII-TCR的参与促进了T细胞和APC之间GJ的形成。实际上作者观察到,与IR小鼠的MHCIIhi型HSC相比,MHCIIhi型HSC中的GJ信号更加活跃(图4B)。GJ是由连接蛋白构建的(Cx)。其中,作者发现Cx43/GJA1在IR小鼠的HSC(尤其是MHCIIhi HSC)和treg中均显著且专一上调(图4C, D)。通过免疫染色,作者还经常观察到IR小鼠BM中HSC和treg之间可能形成基于Cx43的GJs(图4E)。在体外,通过钙黄蛋白(只能通过GJs在细胞之间转移),作者验证了IR小鼠的HSC,特别是MHCIIhi HSC和treg之间建立了功能性GJs(图F, G),并且这种作用是细胞接触和取决于MHCII(图4H)。当IR小鼠BM HSC与敲除Cx43的自体Tregs共培养时,钙黄蛋白转移几乎消失(Cx43-/-;图4H)。同时,Cx43-/- BM Tregs的过继转移与Cx43-精通的BM Tregs相比具有相似的免疫抑制和激活表型,未能逆转Treg耗尽和对HSC衰老的拯救作用(图4I,J)。这些结果表明,衰老的HSC和BM treg之间通过MHCII-TCR的作用建立了基于Cx43的GJs。
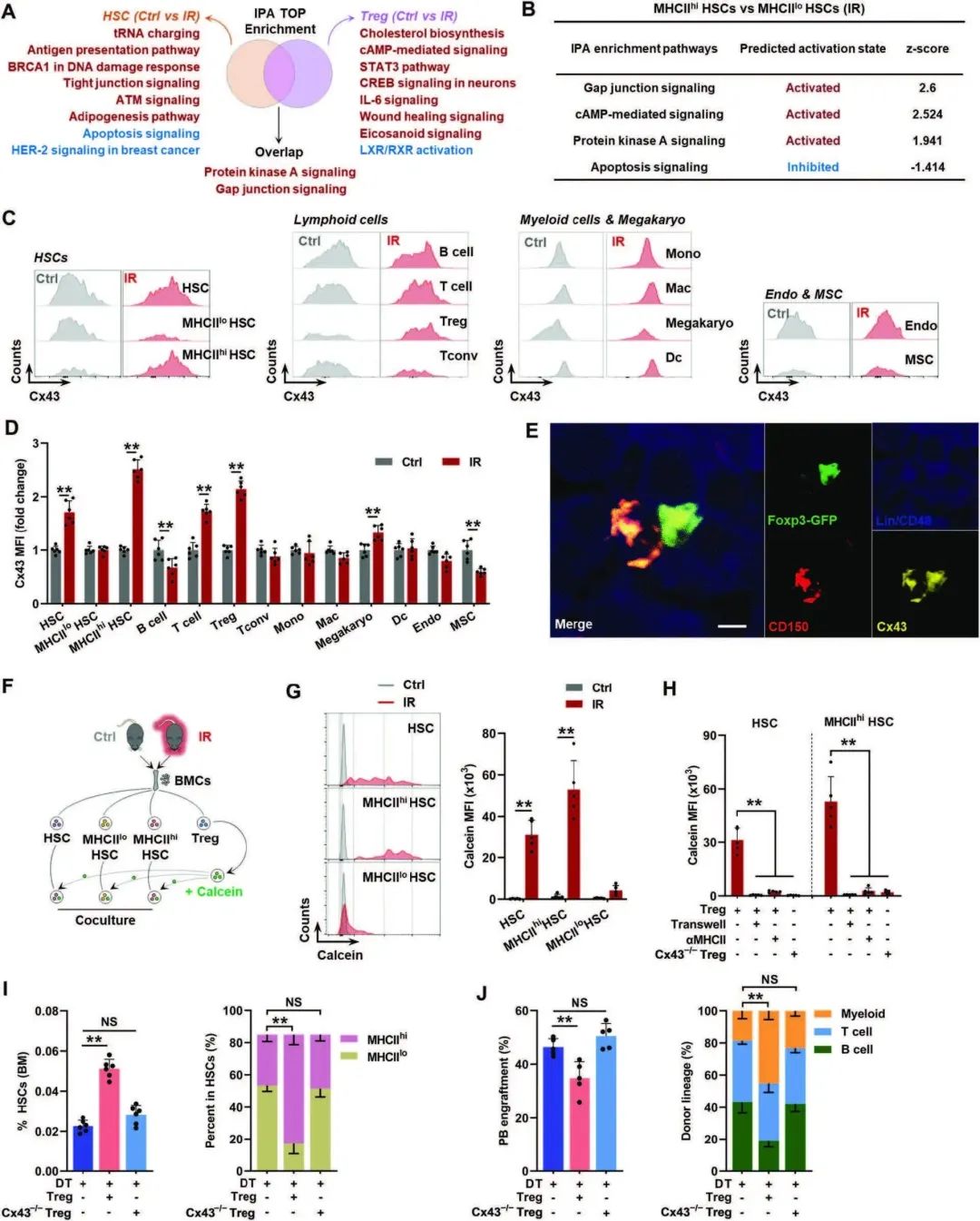
图4
05 BM treg通过GJs转移cAMP,减少衰老hsc的凋亡启动
然后,通过研究老化HSC在用Tregs建立GJs时的结果。在最富集的信号通路中,作者注意到在IR小鼠的HSC中大量存在被抑制的凋亡信号(图4A)。此外,MHCIIhi型HSC中的凋亡信号比MHCIIlo型HSC中的更受抑制(图4B),推断BM-Treg相互作用在HSC凋亡信号传导中发挥作用。为了支持这一发现,尽管IR小鼠MHCIIhi和MHCIIlo造血干细胞中凋亡细胞的比例在稳态下相当(补充图5a),硼烷分析显示,IR小鼠对BIM(图5A)和BID的凋亡启动量远低于MHCIIlo。此外,造血干细胞,特别是MHCIIhi造血干细胞在IR小鼠中的抗凋亡能力是BM-Tregs依赖性的(图5B)。在体外,来自IR小鼠的MHCIIhi HSC的凋亡启动,而不是MHCIIlo HSC,在存在自体BM Tregs的情况下被减弱(图5C),并且这种效应是细胞接触和MHCII依赖的(图5D)。值得注意的是,作者还观察到Cx43对于IR小鼠MHCIIhi HSC的体内抗凋亡是必不可少的(图5B)和离体(图5D),表明GJs的参与。GJs通过小于1200da的可溶性因子的传递调节邻近细胞的胞内信号传导。RNA-seq显示,由次级信使cAMP激活的PKA信号在IR小鼠的BM HSCs和tregs中都被强烈激活(图4A),并且在IR小鼠的MHCIIhi HSC中激活比MHCIIlo HSC中更明显(图4B)。与相关报道一致,作者也发现IR小鼠BM Tregs中cAMP水平的富集和显著升高(图5E)。cAMP-protein kinase A (PKA)信号正调控HSC存活,提示Tregs可能通过GJs将cAMP转移到衰老的HSC中,促进其存活。实际上,作者测量了IR小鼠MHCIIhi HSC中cAMP水平、PKA激酶活性及其下游CREB活性的显著增加(图F-H),并且cAMP-PKA信号的过度激活依赖于BM Tregs中Cx43的表达(图5I)。然而,选择性抑制剂H89对PKA的抑制几乎消除了类似BM Tregs对IR小鼠MHCIIhi HSC的促存活作用(图5J)。为了进一步验证这一假设,作者使用Alexa Fluor 488标记的cAMP (488-cAMP)跟踪了cAMP的转移。如图所示,BM Tregs迅速而显著地将488-cAMP转移到IR小鼠的MHCIIhi HSC而不是MHCIIlo HSC(图5K),并且这种作用依赖于细胞接触、MHCII和Cx43(图5L)。这些数据表明,BM Tregs通过GJs转移cAMP以减少衰老HSC的凋亡启动。
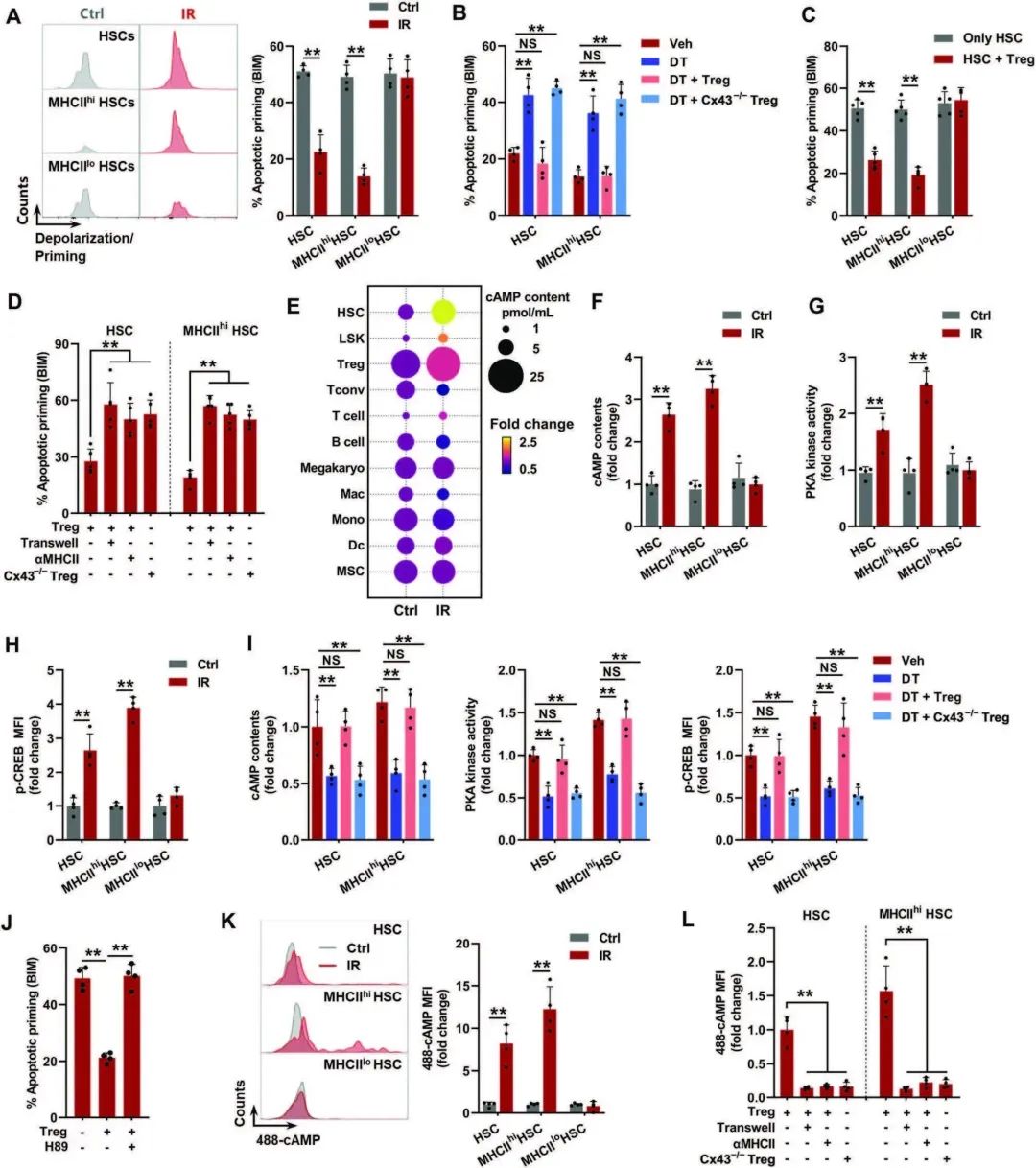
06 HSC-treg双向相互作用与HSC生理老化有关
生理性HSC衰老的另一个特征是DNA突变积累,这种积累在中年很明显。有趣的是,HSC衰老表型(图6A)和BM Tregs的克隆扩增(图6B, C)从中年到老年水平已经很明显,伴随着DNA突变的急剧积累,特别是在MHCIIhi HSC中(图6D)。与IR小鼠类似,作者也发现了中年小鼠HSC室中MHCIIhi HSC的过度代表(图6E)。
因此,与treg的独特邻接性(图6F, G), GJ与Tregs的形成(图6H)和cAMP-PKA信号过度激活(图6I),以及凋亡启动减少(图6J),在中年中年小鼠的造血干细胞中,特别是MHCIIhi中,观察到凋亡启动的减少。这些改变也在转录组学水平上得到了证实(图6K)。在转录组学和功能水平上,中年小鼠的MHCIIhi HSC表现出与老年小鼠相似的衰老特征和谱系偏差(图6L, M)。然而,Treg缺失显著减轻了中年小鼠的HSC衰老表型(图6N-P),证明HSC-treg双向相互作用参与HSC生理性老化。
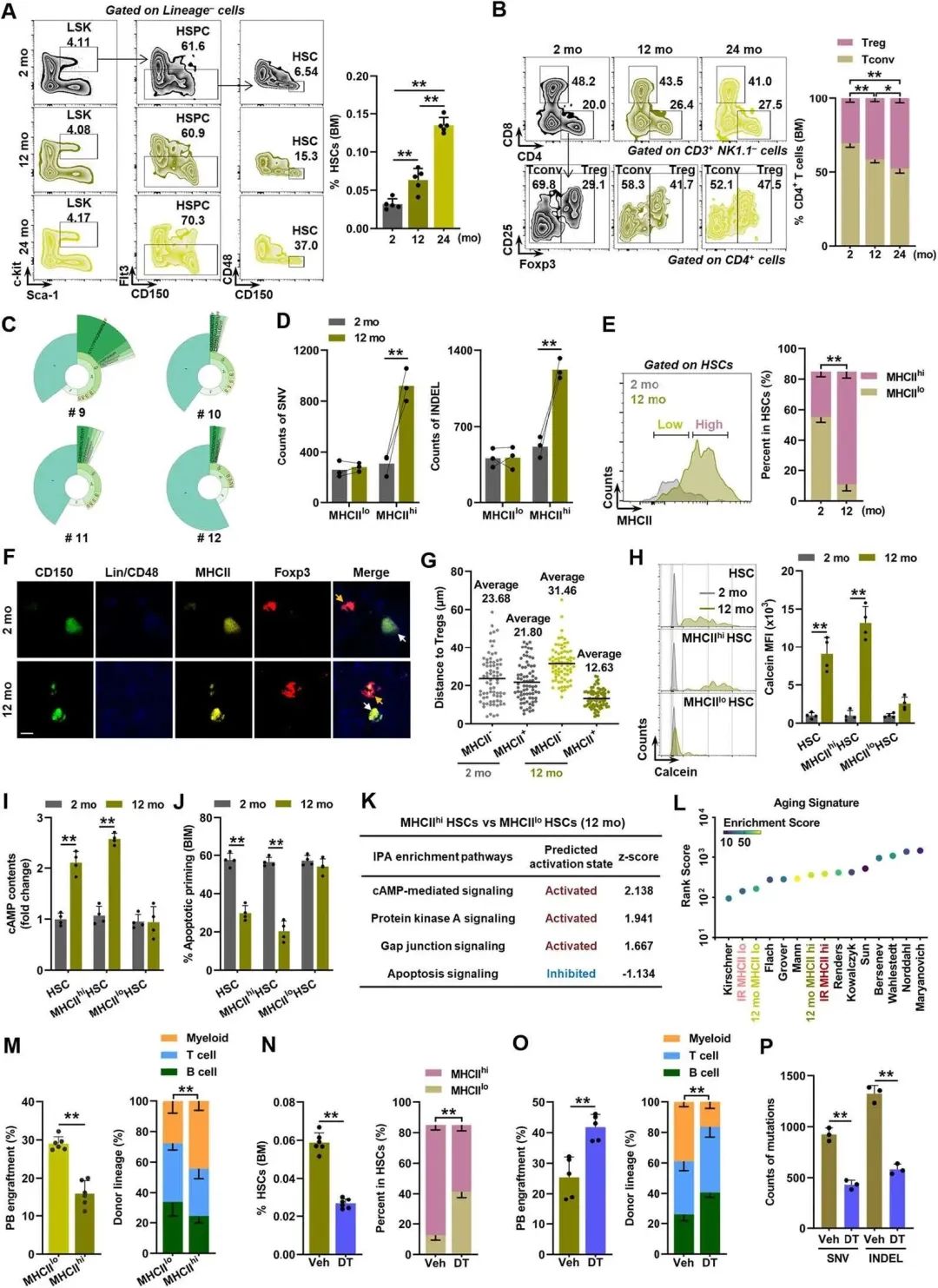
图6
07 通过干预老化HSC – treg相互作用预防HSC衰老
最后,作者评估了研究结果的转化潜力。由于人源抗hCD25抗体(daclizumab)可获得并广泛应用于临床环境,作者首先测试了抗鼠CD25 (αCD25)在HSC衰老中消耗treg的治疗潜力(图7A)。αCD25给药显著减少IR小鼠中BM treg的数量,而不引起CD8+ T细胞的显著活化。同时,αCD25给药后,IR小鼠MHCIIhi HSC扩增明显减弱(图7B),同时显著减轻了DNA突变负荷(图7C、D),抑制了cAMP-PKA信号,增强了HSC的凋亡启动。
最后,αCD25对IR小鼠HSC的衰老表型有明显的改善(图7E)。Gap27是一种肽,在体内已被证明适合干扰基于Cx43的GJs的形成和稳定性。在体外,作者证实Gap27有效阻断了GJ介导的cAMP在IR小鼠BM HSC和treg之间的转移。相应的,GAP27在体内显著缓解了造血干细胞cAMP-PKA信号的激活和凋亡抵抗,从而减轻了IR小鼠造血干细胞的衰老表型(图7B-E)。同样,给药αCD25或GAP27可显著挽救中年小鼠造血干细胞的早衰表型(图7F-J)。因此,系统给药αCD25或GAP27有望预防HSC衰老,至少部分干预了老化的HSC- treg相互作用。
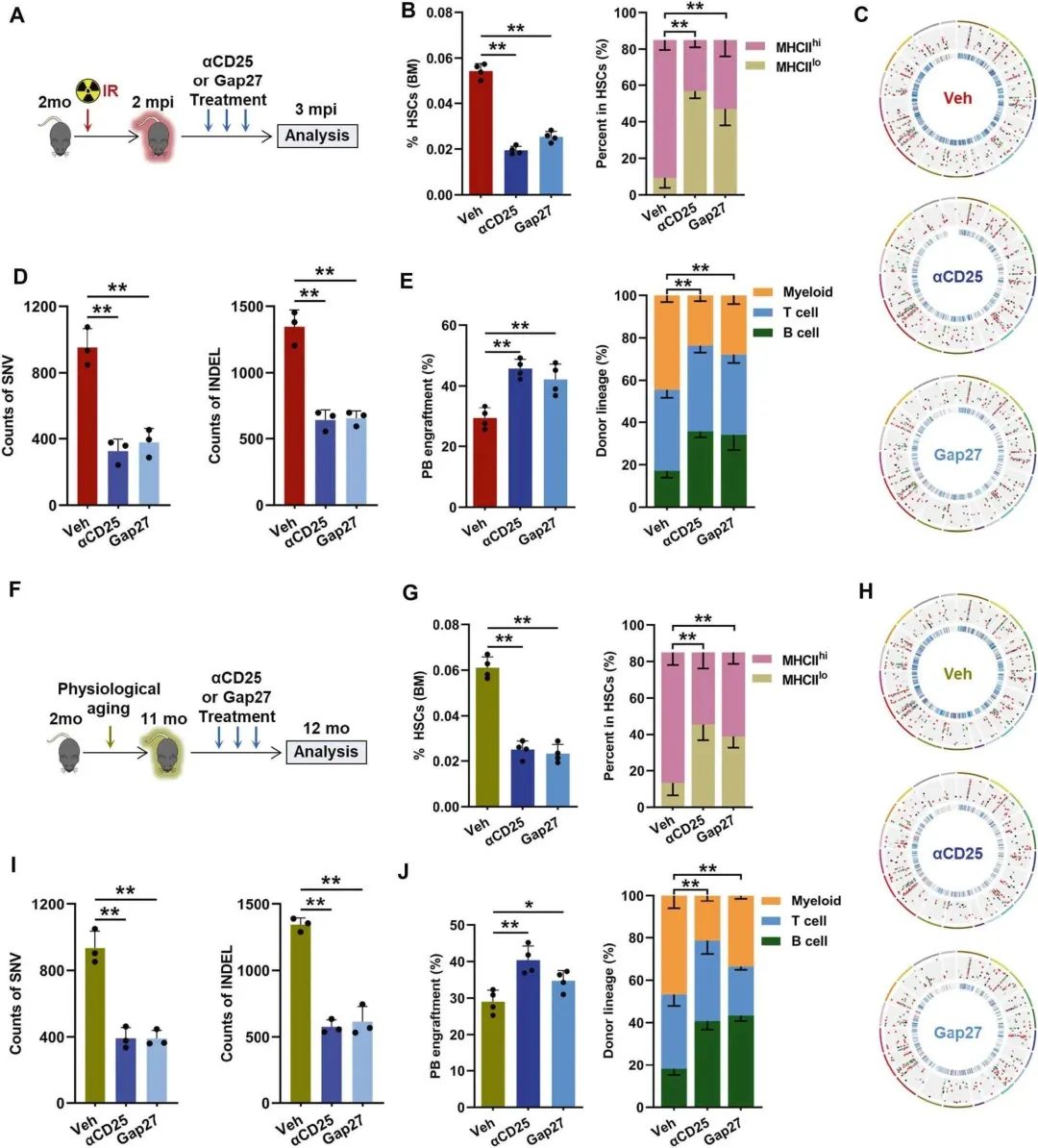
图7
结论
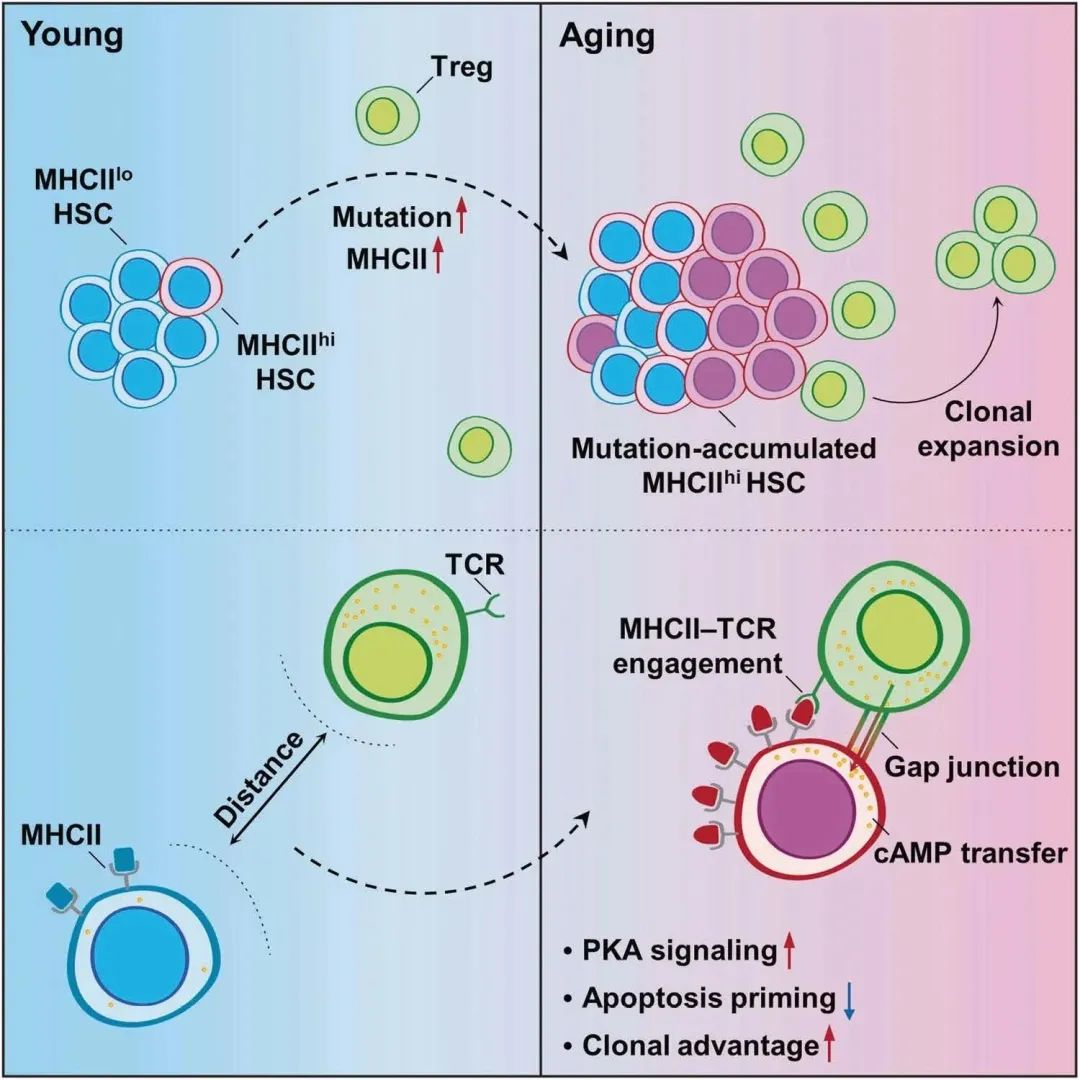
图8
BM Tregs在HSC衰老中的调节作用示意图(图8)特化骨髓(BM)微环境,称为HSC生态位,通过细胞结合或分泌因子或物理相互作用形式的细胞间信号维持HSC池的大小和功能。
总之,本研究凸显免疫细胞及膜蛋白在HSC衰老中的关键作用,为实现提前预防和治疗HSC衰老提供了理论基础和新策略。未来通过系统阐明不同免疫细胞亚群与干细胞的互动,开发靶向干预HSC-Treg轴的新药物,有望抑制组织干细胞的加速衰老,延缓相关疾病的发生。
参考文献
Liao W, Liu C, Yang K, Chen J, Wu Y, Zhang S, Yu K, Wang L, Ran L, Chen M, Chen F, Xu Y, Wang S, Wang F, Zhang Q, Zhao J, Ye L, Du C, Wang J. Aged hematopoietic stem cells entrap regulatory T cells to create a prosurvival microenvironment. Cell Mol Immunol. 2023 Oct;20(10):1216-1231. doi: 10.1038/s41423-023-01072-3. Epub 2023 Aug 30. PMID: 37644165; PMCID: PMC10541885.
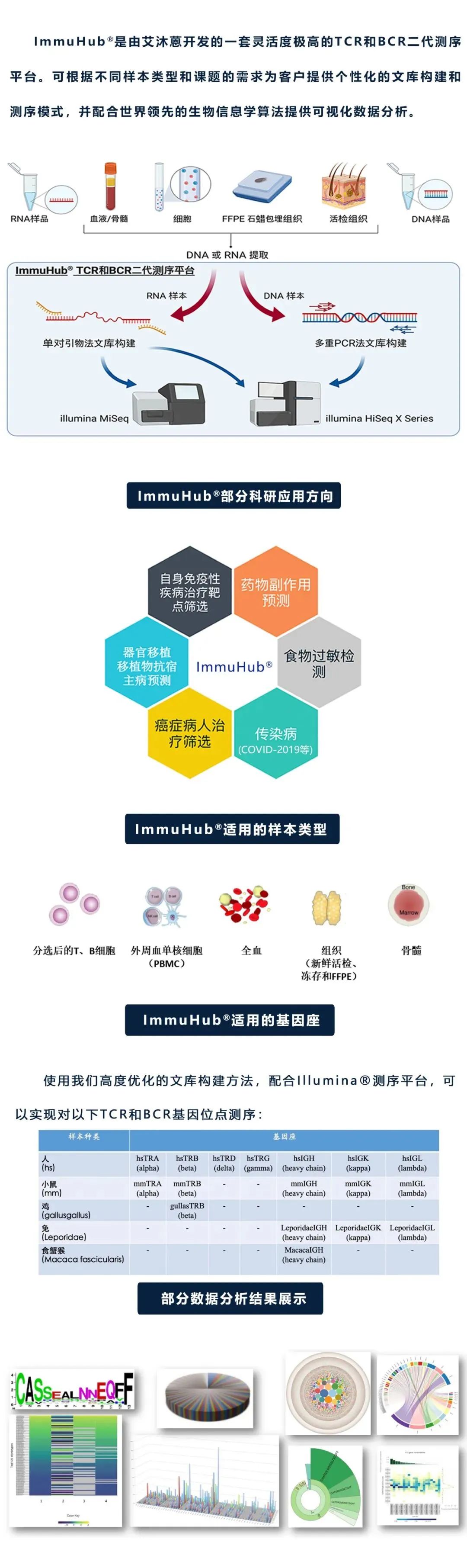

杭州艾沐蒽生物科技有限公司成立于2016年,是国际前沿的专注于免疫驱动医学技术的国家高新技术企业。创始人团队来自美国芝加哥大学,在2010年开始使用免疫组基因高通量测序技术开展各种疾病相关研究,于2016年通过自主研发,全国首家推出NGS-MRD血液肿瘤微小残留病(MRD)检测Seq-MRD®,并授权泛生子(纳斯达克代码:GTH)使用。同时,公司拥有Immun-Traq®肿瘤治疗伴随诊断、Immun-Cheq® |T细胞免疫测评以及ImmuHub®免疫组测序科研服务产品,并布局有基于AI机器学习算法的T-classifier®肿瘤早筛、单细胞测序、TCR-T和抗体发现等平台管线。公司构建几十项发明专利和软件著作权为核心的自主知识产权体系,为医院临床、生命科学研究、新药开发等提供解决方案和产品。艾沐蒽专注于通过解码适应性免疫系统来改变疾病的诊断和治疗,并致力于推进免疫驱动医学领域发展。
